Was ist die Besonderheit der Natur von Viren? Allgemeine Eigenschaften von Viren. Pathogenese viraler Erkrankungen Die Natur von Viren und ihre Untersuchung
Auf der Jahrestagung der American Society of Clinical Oncology, die gerade in San Francisco stattfand, gab es einen aufsehenerregenden Bericht von Wissenschaftlern, die im Gewebe von Prostatakrebspatienten ein Virus entdeckten, das zuvor nur bei krebskranken Mäusen gefunden wurde. Und da das Virus bekannt ist, ist es viel klarer, wie man Antitumormedikamente herstellen kann. Die virale Krebstheorie wurde übrigens bereits in den 1940er Jahren vom sowjetischen Wissenschaftler Lev Zilber begründet.
Darüber hinaus wurde dieses Virus nur im Gewebe von Patienten gefunden, die einen bestimmten genetischen Defekt hatten.
„Wir sagen nicht, dass das Virus die direkte Ursache der Krankheit ist“, sagte der Co-Autor der Studie, Dr. Eric Klein von der Cleveland Clinic. „Aber diese aufregende Entdeckung weist auf völlig neue Wege für die Untersuchung von Prostatakrebs hin.“
Forscher wissen noch nicht, wie das Mausvirus in den menschlichen Körper gelangt, sie vermuten jedoch, dass es genetisch vererbt werden könnte. Dr. Klein und sein Kollege Dr. Joe Derisi von der University of California nutzten die „Genchip“-Technologie. Ein ähnliches Diagnostikum wurde von russischen Wissenschaftlern des Instituts für Molekularbiologie der Russischen Akademie der Wissenschaften erstellt. Auf einer speziellen Platte platzierte Derisi charakteristische Abschnitte des genetischen Materials von 20.000 bekannten Viren. Klein stellte ihm 86 Gewebeproben seiner Prostatakrebspatienten zur Verfügung. Aus ihnen wurden DNA-Proben entnommen und auf einen Chip gegeben. Die DNA der 20 Patienten, bei denen das mutierte Gen nachgewiesen wurde, stimmte mit der DNA des Maus-Onkovirus überein. Eine Mutation ist eine Duplikation eines Gens, das die Produktion von Enzymen kodiert, die in den Körper eindringende Viren zerstören.
Es stellte sich heraus, dass Männer mit einem Doppelgen deutlich weniger dieser Enzyme produzieren. Unter 66 Patienten mit einem normalen Gen wurde das Virus nur bei einem nachgewiesen. Wissenschaftler wollen Hunderte kranke und gesunde Menschen untersuchen, um den Zusammenhang zwischen dem Vorhandensein des Virus und Prostatakrebs zu klären.
Die Entdeckung amerikanischer Wissenschaftler wird zu einer weiteren praktischen Bestätigung der virusogenetischen Krebstheorie, die bereits in den 40er Jahren des 20. Jahrhunderts von Lev Zilber formuliert wurde.
Lev Zilber entwickelte die Theorie über die Entstehung von Krebs im Gefängnis, ein halbes Jahrhundert vor der wissenschaftlichen Welt
Akademiker Lev Kiselev, Sohn von Lev Zilber, beantwortet die Fragen der Iswestija.
Es ist bekannt, dass Lew Alexandrowitsch seine Theorie in den 40er Jahren im Lager entwickelte. Aber die Virus-Hypothese wurde schon einmal geäußert?
Die ersten Hypothesen über die virale Natur von Krebs wurden zu Beginn des 20. Jahrhunderts aufgestellt, unter anderem von unserem Landsmann Ilya Mechnikov. Doch Lev Zilber formulierte eine ganzheitliche virogenetische Theorie, die ihrer Zeit weit voraus war.
- Aber die Zeitgenossen akzeptierten die neue Theorie nicht?
Ja, 20 Jahre lang hat er im Alleingang bewiesen, dass er Recht hatte. Erst in den 60er Jahren erfolgte die erste experimentelle Bestätigung der Theorie. Große Unterstützung leistete die Arbeit des Tschechen Jan Svoboda vom Institut für Genetik der Tschechoslowakei, er lebt noch.
- Gibt es heute keine Zweifel an der Richtigkeit von Lew Alexandrowitsch?
Man geht heute davon aus, dass bis zu 25 % aller Krebserkrankungen unter Beteiligung von Onkoviren entstehen. Dies ist insbesondere bei Leberkrebs, der durch chronische Hepatitis-B- und C-Viren verursacht wird, sowie bei Gebärmutterhalskrebs (humanes Papillomavirus) nachgewiesen. Es gibt Hinweise darauf, dass Krankheiten wie Brustkrebs, Magenkrebs und einige andere nicht ohne die Beteiligung von Viren entstehen. Es ist außerdem erwiesen, dass alle Tumoren bei Tieren durch Viren verursacht werden. Die Worte meines Vaters „Krebs ist eine Erkrankung des Genoms“ erwiesen sich als prophetisch, denn die Entschlüsselung des Genoms war damals noch unglaublich weit entfernt.
Großartiger Wissenschaftler, kluger Mann
Lev Zilber wurde 1894 in Pskow geboren. Nach seinem Abschluss an der medizinischen Fakultät arbeitete er in Moskau und Baku, beteiligte sich an der Ausrottung der Pest in der UdSSR und entwickelte die Theorie der durch Zecken übertragenen Enzephalitis, die darauf hindeutet, dass das Krankheitsvirus durch Zecken übertragen wurde. Er wurde aufgrund absurder Anschuldigungen zweimal inhaftiert (1937–39 und 1940–44). Sein Bruder, der Schriftsteller Veniamin Kaverin, und seine Ex-Frau Sinaida Ermolyeva, bekannt als Erfinderin des „sowjetischen Penicillins“, kämpften selbstlos für seine Freilassung. Es ist bekannt, dass der Bruder als Prototyp für Sanya Grigoriev aus Kaverins Erzählung „Zwei Kapitäne“ diente, die von Millionen Menschen geliebt wird. Der Roman „Offenes Buch“ ist Ermolyeva gewidmet.
Zilber entwickelte seine Theorie über die Entstehung von Krebs im Gefängnis, indem er Experimente in einer wissenschaftlichen „Sharashka“ durchführte. Ratten und Mäuse wurden für ihn von Gefangenen gefangen, bei denen er mit Tabak bezahlte. Zilber untersuchte die Mechanismen der Tumorentstehung und kam zu dem Schluss, dass ein Virus, wenn es in eine gesunde Zelle eindringt, seine genetische Basis verändert, sodass sich die Zelle der Kontrolle des Körpers entzieht und beginnt, sich ungehindert zu teilen – so entsteht ein Tumor. Lew Alexandrowitsch veröffentlichte 1945 den ersten Artikel über seine Theorie in unserer Zeitung. Im selben Jahr erschien seine Monographie zu diesem Thema.
Quelle:
Weitere Artikel in Gesundheit:
| 14. Dezember 2016 |
20. Mai 2016 |
17. Mai 2016 |
| 22. November 2015 |
Viren wurden vom russischen Botaniker D.I. entdeckt. Ivanovsky (1864 – 1920) im Jahr 1892, als er die Mosaikkrankheit von Tabakblättern untersuchte. Der Begriff „Virus“ wurde erstmals 1898 vom niederländischen Wissenschaftler M. Beijerinck (1851 – 1931) vorgeschlagen.
Derzeit sind etwa 3.000 verschiedene Arten von Viren bekannt.
Die Größe der Viren reicht von 15 bis 350 nm (die Länge einiger filamentöser Viren erreicht 3.000 nm; 1 nm = 1·10–9 m), d. h. Die meisten von ihnen sind im Lichtmikroskop (submikroskopisch) nicht sichtbar und ihre Untersuchung wurde erst nach der Erfindung des Elektronenmikroskops möglich.
Im Gegensatz zu allen anderen Organismen haben Viren keine zelluläre Struktur!
Reifes Viruspartikel (d. h. extrazellulär, ruhend – Virion) Die Struktur ist sehr einfach: Sie besteht aus einem oder mehreren Nukleinsäuremolekülen, aus denen sie besteht Kern Virus- und Proteinhülle (Kapsid)- das sind die sogenannten einfache Viren.
Komplexe Viren(Zum Beispiel, Herpes oder Grippe) Darüber hinaus enthalten Kapsidproteine und Nukleinsäure zusätzliche Lipoprotein Membran(Hülle, Superkapsid, gebildet aus der Plasmamembran der Wirtszelle), verschiedene Kohlenhydrate Und Enzyme(Abb. 3.1).
Enzyme erleichtern das Eindringen viraler NK in die Zelle und die Freisetzung der resultierenden Virionen in die Umgebung ( Neuraminidase Myxoviren, ATPase Und Lysozym einige Phagen usw.) und sind auch an den Prozessen der Transkription und Replikation viraler NK beteiligt (verschiedene). Transkriptasen Und Nachbildung).
Proteinhülle schützt die Nukleinsäure vor verschiedenen physikalischen und chemischen Einflüssen und verhindert außerdem das Eindringen zellulärer Enzyme in sie und damit ihren Abbau (Schutzfunktion). Außerdem enthält das Kapsid einen Rezeptor, der zum Rezeptor der infizierten Zelle komplementär ist – Viren infizieren einen genau definierten Bereich von Wirten (definierende Funktion).
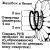
Virionen viele Pflanzenviren und eine Reihe von Phagen haben Spiral- ein Kapsid, in dem die Proteinuntereinheiten (Kapsomere) spiralförmig um eine Achse angeordnet sind. Zum Beispiel VTM ( Tabakmosaikvirus) hat die Form von Stäbchen mit einem Durchmesser von 15–17 nm und einer Länge von bis zu 300 nm (Abb. 3.2.). In seinem Kapsid befindet sich ein Hohlkanal mit einem Durchmesser von 4 nm. Das genetische Material von TMV ist Reis. 3.2.
Die Struktur des Tabakmosaikvirus. In der Rille des helikalen Kapsids befindet sich dicht gepackte einzelsträngige RNA. Für Virionen mit helikalem Kapsid zeichnet sich durch einen hohen Proteingehalt (90 – 98 %) im Verhältnis zu aus
In der Rille des helikalen Kapsids befindet sich dicht gepackte einzelsträngige RNA. Für Virionen mit helikalem Kapsid zeichnet sich durch einen hohen Proteingehalt (90 – 98 %) im Verhältnis zu aus
Die Kapside von Virionen vieler Viren (z. B. Adenovirus, Virus Herpes, gelbes Virus Rübenmosaike– VZhMT) haben die Form eines symmetrischen Polyeders, am häufigsten eines Ikosaeders (ein Polyeder mit 12 Ecken, 20 Dreiecksflächen und 30 Kanten). Man nennt solche Kapside isometrisch(Abb. 3.3.). In solchen Virionen beträgt der Proteingehalt etwa 50 % relativ zu NK.

Ein Virus enthält immer eine Art Nukleinsäure (entweder DNA oder RNA), daher werden alle Viren in DNA-haltige und RNA-haltige Viren unterteilt. Die Nukleinsäuremoleküle im Virion können linear (RNA, DNA) oder ringförmig (DNA) sein. Darüber hinaus können diese Nukleinsäuren aus einer oder zwei Ketten bestehen. Virales NK hat 3 bis 200 Gene.
Die Nukleinsäure des Virus vereint die Funktionen beider Säuren (DNA und RNA) – Speicherung und Übertragung von Erbinformationen sowie Kontrolle der Proteinsynthese.
Im Gegensatz zu Viren enthalten alle Zellorganismen beide Arten von Nukleinsäuren.
Bakterielle Viren haben eine komplexere Struktur – Bakteriophagen(Abb. 3.4.). Sie bestehen aus Kopf und Schwanz (Stab und Hülle, Grundplatte und Fortsatzfilamente). Ein langes NA-Molekül (RNA oder DNA) ist im Inneren des Bakteriophagenkopfes (Proteinhülle) spiralförmig gefaltet.
Zu den Viren zählen auch Viroide– Infektionserreger, bei denen es sich um einzelsträngige zirkuläre RNAs mit niedrigem Molekulargewicht (kurz) handelt, die nicht für ihre eigenen Proteine kodieren (ohne Kapsid). Sie sind Erreger einer Reihe von Krankheiten.
ZU

IN
Reis. 3.4.
Die Struktur eines Bakteriophagen. Threads des Prozesses
Wissenschaftler gehen davon aus, dass Viren vor etwa 3 Milliarden Jahren aus den Nukleinsäuren von Organismen entstanden sind (Prokaryoten) als Ergebnis der Isolierung freier Fragmente aus dem Genom, die die Fähigkeit erworben haben, eine Proteinhülle zu synthetisieren und sich innerhalb von Zellen zu teilen (verdoppeln, replizieren). Es wird angenommen, dass immer noch neue Arten von Viren aus dem Genom von Bakterien und Bakterien gebildet werden Eukaryoten(Kerne, Plastiden, Mitochondrien) .
In der Natur sind Viren von großer Bedeutung, da sie allgegenwärtig sind und alle Gruppen lebender Organismen befallen und häufig verschiedene Krankheiten verursachen.
Es sind mehr als 1000 Krankheiten bekannt Pflanzen verursacht durch Viren (RNA-haltig). Die häufigsten sind vielfältig Nekrose(Bereiche mit abgestorbenem Gewebe), Mosaik(Flecken, Flecken, Streifen auf Pflanzenorganen), bei denen Parenchymgewebe beschädigt wird, die Anzahl der Chloroplasten abnimmt, Phloem zerstört wird usw.; Es werden faltige oder verkümmerte Blätter beobachtet. Viren verursachen ein verkümmertes Pflanzenwachstum, was zu geringeren Erträgen führt.
VZhMT – Rübengelbmosaikvirus, VTM – Tabakmosaikvirus, VKKT – Tomaten-Zwergbuschvirus.
Das Auftreten von Streifen auf den Blüten einiger Tulpensorten (bunt) wird ebenfalls durch einen Virus verursacht, aber Blumenzüchter verkaufen diese Tulpen und geben sie als besondere Sorte aus.
U Tiere Viren (DNA und RNA enthaltend) verursachen Krankheiten wie: Maul-und Klauenseuche(bei Rindern) Tollwut(bei Hunden, Füchsen, Wölfen), Myxomatose(bei Ratten) Sarkom, Leukämie Und Pest(bei Hühnern) usw. Sehr häufig infizieren sich Menschen mit diesen Krankheiten (durch Kontakt mit infizierten Tieren).
U Person Viren verursachen Krankheiten wie: Pocken(Variola-Virus) Schweinchen(Paramyxovirus), Grippe(Myxovirus), Atemwege Krankheiten(ARI; Rhinoviren RNA-), infektiöse Hepatitis, Polio(Kinderlähmung; Picornavirus), Tollwut, Herpes, AIDS(Humanes Immundefizienzvirus – HIV).
Grippe - die einzige Infektionskrankheit, die sich in Form periodischer globaler Epidemien manifestiert, die für das menschliche Leben gefährlich sind. Die infektiösen Eigenschaften des Influenzavirus (betrifft die Schleimhäute der Atemwege) hängen wie bei anderen Viren von bestimmten Proteinen der Virushülle ab, die sich durch Rekombinationen oder Mutationen ständig verändern. Daher verursachen neue Stämme des Influenzavirus neue Epidemien, da der Mensch noch keine Immunität gegen sie entwickelt hat.
So wurden im Winter 1968/69 in den USA 50 Millionen Fälle registriert Hongkong-Grippe während 70.000 Menschen starben. Die Epidemie von 1918/19 erfasste den gesamten Globus, verlief in drei Wellen und forderte 20 Millionen Menschenleben.
Viruserkrankungen sind schwer zu behandeln, da Viren nicht empfindlich auf Antibiotika reagieren. Glücklicherweise verhindert das Immunsystem in vielen Fällen, dass sich die Infektion weiter ausbreitet.
Zahlreiche Viruserkrankungen bei Mensch und Tier können dadurch verhindert werden Immunisierung– Durchführung vorbeugender Impfungen, die es Ihnen ermöglichen, eine Immunität gegen Viren zu entwickeln.
Viren werden vom Menschen häufig in der mikrobiologischen Forschung (Biotechnologie, Gentechnik) eingesetzt. Es ist möglich, Viren zur Bekämpfung von Pflanzenschädlingen einzusetzen.
In den USA mit Baumwollkapsel effektiv mit Hilfe des Virus bekämpfen. Diese Bekämpfungsmethode ist praktisch harmlos – das Virus ist in der Regel artspezifisch (d. h. es befällt nur einen bestimmten Organismustyp).
Es wurde auch festgestellt, dass z. B. Reis-nekrotisches Mosaikvirus hemmt das Wachstum von Reis. Aber auch andere Pflanzen, zum Beispiel Jute(eine Quelle grober Fasern für Taschen und Seile), wachsen besser, wenn sie von diesem Virus befallen sind, als wenn sie gesund sind. Wissenschaftler können dieses Phänomen noch nicht erklären.
Bakteriophagen infizieren Bakterien (dringen in sie ein und zerstören sie aktiv), darunter auch pathogene. Daher ist es möglich, sie zur Vorbeugung und Behandlung vieler Infektionskrankheiten sowie zur Bekämpfung pathogener Bakterien einzusetzen: Pest, Typhus, Cholera usw.
Vorlesung 1
Natur, Ursprung und Struktur von Viren.
Modul 1
Integrierter Zweck des Moduls
Das umfassende Ziel des Moduls besteht darin, den Studierenden ein Verständnis für die grundlegenden Eigenschaften von Viren zu vermitteln: ihre Natur, Herkunft, Architektur und Morphologie viraler Partikel, Symmetrietypen und chemische Eigenschaften von Viren. Diese Informationen sollen eine theoretische Grundlage für ein weiteres Verständnis des biologischen Wesens von Viren sein, das sich in den Prozessen der Reproduktion und viralen Pathogenese ausdrückt. Das Modul besteht aus drei Vorlesungen, deren Stoff es Ihnen ermöglicht, das gesetzte Ziel zu lösen.
NATUR DER VIREN
Seit der Entdeckung der Viren bis heute haben sich die Vorstellungen über die Natur von Viren erheblich verändert.
D. I. Ivanovsky und andere Forscher dieser Zeit betonten zwei Eigenschaften von Viren, die es ermöglichten, sie aus der Gesamtmasse der Mikroorganismen zu isolieren: Filtrierbarkeit und Unfähigkeit, sich in allen künstlichen Nährmedien zu vermehren. Später stellte sich heraus, dass diese Eigenschaften nicht absolut sind, da filtrierbare Formen von Bakterien und Mykoplasmen entdeckt wurden, die auf künstlichen Nährmedien wachsen und in ihrer Größe den größten Viren (menschliche und tierische Pockenviren) nahekommen.
Selbst wenn jedoch keine Integration stattfindet und sich das Virusgenom in einem autonomen Zustand befindet, ist das Auftreten einer Infektion auf die Konkurrenz zwischen dem Virus- und dem Zellgenom zurückzuführen.
Zu den einzigartigen Eigenschaften des Virus gehört seine Fortpflanzungsmethode, die sich stark von den Fortpflanzungsmethoden aller anderen Zellen und Organismen unterscheidet (binäre Spaltung, Knospung, Sporenbildung). Viren wachsen nicht und ihre Reproduktion wird als disjunktive (getrennte) Reproduktion bezeichnet, was die Uneinheitlichkeit im Raum (auf dem Territorium der Zelle) und im Zeitpunkt der Synthese viraler Komponenten (Nukleinsäuren und Proteine) mit anschließender Montage und Bildung betont von Virionen.
Im Zusammenhang mit dem oben Gesagten kam es immer wieder zu Diskussionen darüber, was Viren sind – lebend oder nichtlebend, Organismen oder Nichtorganismen. Natürlich haben Viren die grundlegenden Eigenschaften aller anderen Lebensformen – die Fähigkeit zur Fortpflanzung, Vererbung, Variabilität, Anpassungsfähigkeit an Umweltbedingungen; Sie besetzen eine gewisse ökologische Nische, sie unterliegen den Evolutionsgesetzen der organischen Welt auf der Erde. Daher entwickelte sich bereits Mitte der 40er Jahre die Vorstellung von Viren als den einfachsten Mikroorganismen. Die logische Weiterentwicklung dieser Ansichten war die Einführung des Begriffs „Virion“, der ein extrazelluläres virales Individuum bezeichnete. Mit der Entwicklung der Forschung zur Molekularbiologie von Viren begannen sich jedoch Fakten zu häufen, die der Vorstellung von Viren als Organismen widersprachen.
Das Fehlen eigener Proteinsynthesesysteme, eine disjunktive Reproduktionsweise, die Integration in das zelluläre Genom, die Existenz von Satellitenviren und defekten Viren, die Phänomene der mehrfachen Reaktivierung und Komplementierung – all das passt nicht gut in die Idee von Viren als Organismen. Diese Idee verliert noch mehr an Bedeutung, wenn wir uns virusähnlichen Strukturen zuwenden – Plasmiden, Viroiden und Erregern wie dem Scrapie-Erreger.
Plasmide (andere Namen sind Episomen, Epiviren) sind doppelsträngige zirkuläre DNA mit einem Molekulargewicht von mehreren Millionen, die von der Zelle repliziert werden. Sie wurden erstmals in Prokaryoten entdeckt und ihre Existenz wird mit verschiedenen Eigenschaften von Bakterien in Verbindung gebracht, beispielsweise mit der Resistenz gegen Antibiotika. Da Plasmide normalerweise nicht mit dem Bakterienchromosom assoziiert sind (obwohl viele zur Integration fähig sind), gelten sie als extrachromosomale Vererbungsfaktoren.
Plasmide wurden auch in Eukaryoten (Hefen und anderen Pilzen) entdeckt; darüber hinaus können gewöhnliche Viren höherer Tiere auch in Form von Plasmiden, also zirkulärer DNA, ohne eigene Proteine, vorkommen und durch zelluläre DNA-Syntheseenzyme repliziert werden. Insbesondere bovine Papillomaviren und das Affenvirus 40 (SV-40) können in Form von Plasmiden existieren. Wenn das Herpesvirus in der Zellkultur persistiert, können Plasmide gebildet werden – zirkuläre DNA, die nur einen Teil des Genoms dieses Virus ausmacht.
Mit Viren verwandt sind Viroide, 1972 von T. O. Diner entdeckte Erreger, die bei einigen Pflanzen Krankheiten verursachen und wie gewöhnliche infektiöse Viren übertragen werden können. Bei ihrer Untersuchung stellte sich heraus, dass es sich um relativ kleine Moleküle zirkulärer superspiralisierter RNA handelte, die aus wenigen, 300–400 Nukleotiden bestanden. Der Mechanismus der Viroidreplikation ist nicht ganz klar.
Abschließend ist noch der Erreger Scrapie zu erwähnen, der Erreger der subakuten transmissiblen spongiformen Enzephalopathie bei Schafen. Wahrscheinlich verursachen ähnliche Wirkstoffe bei Tieren und Menschen andere Formen der spongiformen Enzephalopathie, die auf der fortschreitenden Zerstörung von Nervenzellen beruhen, wodurch das Gehirn eine spongiforme (spongioforme) Struktur erhält. Der Scrapie-Erreger ist proteinartig und hat sogar einen besonderen Namen erhalten – Prion (von den Wörtern proteinaceous infectious Particle – Protein-infektiöser Partikel). Es wird angenommen, dass dieses Protein sowohl ein Induktor als auch ein Produkt eines zellulären Gens ist, das autonom geworden ist und sich der Regulierung entzogen hat („verrückt geworden“).
Alle Viren, einschließlich Satelliten- und Defektviren, Plasmide, Viroide und sogar Scrapie-Erreger (ihre Gene), haben etwas gemeinsam, das sie verbindet. Bei allen handelt es sich um autonome genetische Strukturen, die in den Zellen von Tieren, Pflanzen, Protozoen, Pilzen und Bakterien, die dafür anfällig sind, funktionieren und sich vermehren können. Anscheinend ist dies die allgemeinste Definition, die es uns ermöglicht, das Reich der Viren zu skizzieren. Basierend auf der formulierten Definition sind Viren zwar keine Organismen, aber dennoch eine einzigartige Lebensform und gehorchen daher den Evolutionsgesetzen der organischen Welt auf der Erde.
URSPRUNG VON VIREN
Über den Ursprung von Viren gibt es unterschiedliche Annahmen. Einige Autoren glaubten, dass Viren das Ergebnis einer extremen Manifestation der regressiven Evolution von Bakterien oder anderen einzelligen Organismen seien. Die Hypothese der regressiven Evolution kann die Vielfalt des genetischen Materials in Viren, ihre nichtzelluläre Organisation, ihre disjunktive Art der Fortpflanzung und das Fehlen proteinsynthetisierender Systeme nicht erklären. Daher hat diese Hypothese derzeit eher historische Bedeutung und wird von der Mehrheit der Virologen nicht geteilt.
Nach der zweiten Hypothese sind Viren die Nachkommen alter, präzellulärer Lebensformen – Protobionten, die der Entstehung zellulärer Lebensformen vorausgingen, von denen aus die biologische Evolution begann. Auch diese Hypothese wird derzeit von der Mehrheit der Virologen nicht geteilt, da sie nicht die gleichen Probleme erklärt, die die erste Hypothese nicht lösen konnte.
Eine dritte Hypothese besagt, dass sich Viren aus genetischen Elementen autonom gewordener Zellen entwickelt haben, obwohl nicht klar ist, welche dieser Elemente zu einer so großen Vielfalt an genetischem Material in Viren geführt haben. Diese Hypothese, die ironischerweise als Hypothese der „laufenden Gene“ bezeichnet wurde, findet die meisten Befürworter, jedoch nicht in der ursprünglichen Form, in der sie ausgedrückt wurde, da sie das Vorhandensein von Formen genetischen Materials in Viren (einzelne Gene) nicht erklärt. (strängige DNA, doppelsträngige RNA), Fehlen in Zellen, Kapsidbildung, Existenz zweier Symmetrieformen usw.
Es ist wahrscheinlich, dass Viren tatsächlich Derivate der genetischen Elemente von Zellen sind, aber sie entstanden und entwickelten sich zusammen mit der Entstehung und Entwicklung zellulärer Lebensformen. Die Natur hat sozusagen alle möglichen Formen des genetischen Materials (verschiedene Arten von RNA und DNA) an Viren ausprobiert, bevor sie sich schließlich für ihre kanonische Form entschieden hat – doppelsträngige DNA, die allen zellulären Formen von Organismen gemeinsam ist, von Bakterien bis hin zu Menschen. Da es sich bei Viren einerseits um autonome genetische Strukturen handelt, andererseits aber nicht in der Lage ist, sich außerhalb von Zellen zu entwickeln, haben sie im Laufe der Jahrmilliarden der biologischen Evolution so unterschiedliche Entwicklungspfade eingeschlagen, dass ihre einzelnen Gruppen keinen sukzessiven Zusammenhang haben gegenseitig. Anscheinend sind verschiedene Gruppen von Viren zu historisch unterschiedlichen Zeiten aus unterschiedlichen genetischen Elementen von Zellen entstanden und daher haben die derzeit existierenden verschiedenen Gruppen von Viren einen polyphyletischen Ursprung, das heißt, sie haben keinen einzigen gemeinsamen Vorfahren. Allerdings gilt die Universalität des genetischen Codes auch für Viren und weist damit darauf hin, dass auch sie ein Produkt der organischen Welt der Erde sind.
ROLLE VON VIREN IN DER EVOLUTION
Da Viren als besondere Lebensformen keine Mikroorganismen sind, ist die Virologie kein Teilgebiet der Mikrobiologie, sondern eine eigenständige wissenschaftliche Disziplin mit eigenem Untersuchungsgegenstand und eigenen Forschungsmethoden.
Natur und Ursprung von Viren
Antigene Variabilität des Influenzavirus und Aspekte seiner Untersuchung.
Die Lösung zur Gewinnung wirksamer abgeschwächter Varianten des Influenzavirus wird durch die einzigartige Plastizität und Variabilität seiner antigenen Eigenschaften erschwert. Fast jährliche Influenza-Epidemien in unterschiedlichen Abständen werden global. In den letzten Jahren ist die Infektion, die Pandemien verursacht, das Influenza-A-Virus. Die Analyse der Antigenverschiebungen innerhalb jedes seiner drei Typen zeigt, dass die Änderung der Antigenzusammensetzung der Virusstämme vom Typ AO zu Typ A allmählich erfolgte und der Übergang vom Typ A1 zu A2 war scharf.
Nachdem 1957 festgestellt wurde, dass ein neuer serologischer Typ A2 in der Natur aufgetaucht war, schien dieser für einige Zeit stabil zu sein, obwohl es geringfügige Veränderungen gab. Doch bereits 1964 erlangten sie Bedeutung, und nach der Epidemie in Hongkong unterschieden sich die Viren so stark, dass man von der Entstehung eines neuen Antigentyps A ausging. Während der Evolution des Virus veränderten sich nicht nur die antigenen Eigenschaften von Oberflächenproteinen , aber auch andere Eigenschaften. Der während der Epidemie von 1971-1972 isolierte Influenzavirusstamm erhöhte im Gegensatz zu zuvor zirkulierenden Stämmen seine Reproduktions- und Neuraminidaseaktivität deutlich, erhöhte die Thermostabilität von Viren stark und erlangte die Fähigkeit, bei Mäusen regelmäßig Virämie zu verursachen (Zakstelskaya et al., 1969). ; Sokolov, Podchernyaeva, 1975).
Während früher Typ-B-Viren relativ stabil waren, gibt es seit 1967 einen kontinuierlichen Wandel (Seihachiro, Mitsuo, 1974). Im April–Mai 1974 wurden neue Stämme des Influenzavirus isoliert, von denen B/Hong Kong 15/72 als neue Variante und andere als Mittelstufe zwischen den alten und neuen Stämmen gelten, was uns eine Neubewertung der Daten ermöglicht die Antigenstabilität des Influenza-B-Virus und deuten auf die Entstehung eines neuen Serotyps hin.
Dadurch entsteht ein Bild signifikanter Antigenveränderungen innerhalb der Typen A und B. Dies erregt natürlich die große Aufmerksamkeit von Wissenschaftlern, die sich mit dem Problem der Influenza befassen. Da selbst das Vorhandensein einer starken Immunität in der Bevölkerung keine so großen antigenen Veränderungen im Influenzavirus verursachen kann, wurde eine Hypothese über die Periodizität von Rekombinationen aufgestellt, die zwischen menschlichen und tierischen Influenzaviren sowie zwischen menschlichen und Vogelviren auftreten. Bei der Entwicklung dieser Hypothese stellte sich heraus, dass Influenza auf natürliche Weise Schweine, Pferde, Truthähne, Hühner, Enten und Seeschwalben befällt, und diese Liste wird immer länger. Sie haben Antikörper gegen menschliche Influenzaviren in ihrem Blutserum. Dies ist nicht überraschend, da die Fragmentierung des Influenzavirus-Genoms nicht nur die Möglichkeit einer intraspezifischen, sondern auch interspezifischen Rekombination bestimmt.
Die präparative Trennung von Neuraminidase und Hämagglutinin eröffnet Perspektiven für eine eingehende Analyse des Zusammenhangs einzelner Merkmale. Einige Forscher (Webster u. a., 1973; Gorev et al., 1974) stellen fest, dass das rekombinante Virus gleichzeitig mit Hämagglutinin Virulenz erlangt. Es gibt eine Reihe von Rekombinanten mit spezifiziertem Hämagglutinin und Neuraminidase.
Derzeit untersuchen viele Virologielabore auf der ganzen Welt Grippeseuchen und analysieren antigenische Beziehungen zur menschlichen Grippe. Die Arbeit in dieser Richtung wird von der WHO koordiniert und gefördert. Die Komplexität dieses Problems erfordert einen mehrdeutigen Lösungsansatz. Parallele Suchen nach anderen Lösungsansätzen für dieses Problem sollten nicht als Alternative betrachtet werden.
In den 40er und 50er Jahren wurde die Entstehung antigener Varianten während der Passage des Virus im Körper immunisierter Tiere experimentell nachgewiesen (Archetti, Horsfoll, 1960). Diese Veränderungen waren ziemlich hartnäckig, da die Viren ihre neue Antigenspezifität bei aufeinanderfolgenden Passagen in ovo und in Abwesenheit von Immunserum beibehielten. Darüber hinaus verändern langfristige Passagen des Influenzavirus durch den Körper nicht immunisierter gesunder Tiere seine biologischen Eigenschaften. Beispielsweise erhielt K. Paucker (1960) bei Passagen des PR8-Stamms lange Zeit ein Virus, das sich antigenisch vom Original unterschied und anderen Arten von Influenzaviren nicht ähnelte. Der Autor geht davon aus, dass zwischen den Passagen 103 und 107 ein Mutant entstanden ist, der anschließend das ursprüngliche Virus ersetzte. Ähnliche Daten werden von K. Zgozelska et al. (1973) bereitgestellt.
Hier sehen wir eine Manifestation des Grundgesetzes der Entwicklung jeder Population, auch einer viralen – der Genpool einer Population verändert sich im Laufe der Zeit: Einerseits wird er durch das Aussterben von Organismen, die einzelne Gene enthalten, erschöpft, und andererseits wird es durch Mutationen angereichert, die zur Entstehung neuer Gene führen.
Die Arbeit von S. Fazekas de Sent Groth, C. Hannoun (1973) über die Selektion spontaner antigener Mutanten des Influenza-A-Virus unter „Immunopress“ (d. h. in Gegenwart von Immunserum) ermöglichte es, die hierarchische Reihenfolge von zu reproduzieren Viren innerhalb jedes Typs. Darüber hinaus stützte er sich bei allen seinen Schlussfolgerungen auf die Indikatoren des Querschnitts-RZGA. In Experimenten zur Selektion von Spätmutanten, die mit Hilfe von Antikörpern gewonnen wurden, gelang es ihm, den natürlichen Prozess der Selektion epidemischer Stämme nachzubilden. Er schlug auch ein einfaches Modell der Wechselwirkung eines Antikörpers mit einem Antigen vor. Der Autor präsentierte die Antigenzone der Proteinhülle des Virus in Form einer kleinen Anzahl von Aminosäure-Proteinketten, die über die Oberfläche des Virus hinausragen. Schematisch sieht es aus wie eine Gabel mit Zähnen unterschiedlicher Länge und Breite, und die entsprechenden Antikörper sind Hohlräume, die zu einigen oder allen Zähnen komplementär sind. Somit führt der Kontakt des Antiserums mit einem verwandten Antigen zur Eliminierung homologer Antigene und Antigene mit nichtkomplementären Regionen, also Mutanten, verbleiben in der Population.
Dieses Diagramm stellt die logische Weiterentwicklung der Grundprinzipien der Immunologie dar, die in den 40er Jahren entstanden sind, über die Wechselwirkung von Antigen und Antikörper und die Theorie der Antikörperbiosynthese. Diesen Arbeiten zufolge weist die aktive Gruppe von Antikörpern zusätzlich zur Konfiguration der bestimmenden Gruppe des Antigens eine Konfiguration auf. Es wurde angenommen, dass diese Gruppen zueinander in einer Beziehung stehen wie ein Objekt zu seinem Spiegelbild. K. Landsteiner (1946) führte Experimente mit einem künstlichen Antigen durch, das durch Komplexierung von Proteinmolekülen mit verschiedenen niedermolekularen Verbindungen gewonnen wurde. Dabei zeigte sich, dass die Spezifität dieses Antigens nur durch eine kleine, an das Protein gebundene Gruppe bestimmt werden kann. Antikörper „erkennen“ ein Antigen nicht, wenn es sich nur in der Position der Methylgruppe im aromatischen Ring von derjenigen, die die Bildung dieser Antikörper stimuliert hat, oder in der räumlichen Position der Hydroxylgruppe unterscheidet (Boyd 1969).
Wenn wir also auf die Frage der Antigenvariabilität des Virus zurückkommen, können wir die Selektionsrolle von Antikörpern in diesem Prozess darlegen. Wie mutierte Partikel in einer Viruspopulation entstehen, ist eine der Fragen, die beantwortet werden müssen, um die Evolution von Influenzaviren zu verstehen.
Jede Viruspopulation enthält spontane Mutanten, die durch die Einwirkung externer oder interner Faktoren entstehen. Abhängig von den erworbenen Eigenschaften kann ein Mutant einen Vorteil bei der Fortpflanzung haben und in der Population dominieren. In einigen Fällen ist es möglich, den Faktor zu identifizieren, der bei der Entstehung des Mutanten eine entscheidende Rolle gespielt hat. Die Pandemie von 1918 ist für Forscher, die sich mit dem Problem der Grippe befassen, von größtem Interesse, da ihr Virus für den Menschen äußerst pathogen war. Eine retrospektive Analyse dieses Virus hat einige Forscher zu der Annahme veranlasst, dass die Pandemie durch ein 1930 isoliertes Schweinegrippevirus verursacht wurde, da die Stämme Antigene aufweisen, die mit denen von Schweineviren verwandt sind. Einer anderen Sichtweise zufolge wird die Zunahme der Virusaktivität durch das Auftreten mutierter Partikel unter dem Einfluss von Senfgas verursacht, das während des Ersten Weltkriegs, also vor der pandemischen Grippewelle, eingesetzt wurde (Blashkovich 1966). Tatsächlich ist Senfgas ein extrem starker biologisch aktiver chemischer Wirkstoff. Seine mutagene Aktivität wurde erstmals von C. Auerbach und T. M. Robson (1946) nachgewiesen. Gleichzeitig wurde festgestellt, dass Senfgas eine direkte mutagene Wirkung auf Chromosomen hat. Später wurde entdeckt, dass Senfgas Mutationen bei Viren und Bakterien verursachen kann. Folglich kann seine mögliche Rolle als mutagenes Mittel nicht ausgeschlossen werden, wenn man berücksichtigt, dass chemische und physikalische Faktoren in allen Entwicklungsstadien genetische Veränderungen in biologischen Objekten verursachen können und Viren offenbar keine Ausnahme bilden.
Zu den Faktoren, die unter natürlichen Bedingungen mutagen wirken können, gehören pharmakologische Arzneimittel. Es gibt Arbeiten, die den Zusammenhang zwischen teratogener Aktivität und der chemischen Struktur von Arzneimittelmolekülen analysieren; Bei Mikroorganismen wird ein ähnliches Phänomen der weit verbreiteten Entstehung arzneimittelresistenter Mutantenformen beobachtet. Auf dem Höhepunkt der Grippe, wenn sich das Virus im Körper vermehrt, nehmen die Patienten Medikamente ein, bei denen es sich um synthetische chemische Verbindungen handelt.
Es ist bekannt, dass antivirale Wirkstoffe nur dann recht wirksam sind, wenn sie die Synthese von Nukleinsäuren selektiv unterdrücken können, also in direkten Kontakt mit dem genetischen Apparat kommen. Offenbar wird aufgrund der Besonderheiten des Influenzavirus-Genoms die Grenze zwischen der rein antiviralen und mutagenen Wirkung chemischer Verbindungen leicht überschritten.
Unsere Experimente zur Untersuchung der Wirkung chemischer Verbindungen auf die Antigenspezifität von Influenzaviren des AO-Serotyps zeigten, dass einige Verbindungen aus der Klasse der Supermutagene Veränderungen verursachen können, die nicht über den homologen Serotyp hinausgehen. Insbesondere die ersten beiden Vertreter der Nitrosoalkylharnstoffe induzierten Mutationen für dieses Merkmal (Chulanova, 1968; Akhmatullina et al. 1974). Wir haben unsere vorgeschlagene Modifikation des RZGA verwendet, die es ermöglichte, den Koeffizienten Ap zu ermitteln und darauf basierend den Grad des Unterschieds in der Antigenspezifität von Wild- und Mutantenviren zu bestimmen.
Experimente mit einer großen Anzahl chemischer Verbindungen brachten einen weiteren Wirkstoff zu Tage: 1,4-Bisdiazoacetylbutan, der aufgrund der Antigenspezifität bei Mutationen aktiv ist. Wir verwendeten auch die Immunopress-Methode; nach der Exposition gegenüber dem Mutagen wurde das Virus in Gegenwart von homologem Serum passagiert. Unmodifizierte Viruspartikel wurden mit komplementären Antikörpern neutralisiert und selektive Bedingungen für induzierte Mutanten geschaffen. Die resultierenden Antigenmutanten wurden in einem Kreuzreaktionstest mit Serum auf Wild- und Mutantenviren sowie in einer Präzipitationsreaktion untersucht und zeigten signifikante Antigenverschiebungen.
Daher wird eine weitere experimentelle Untersuchung induzierter Mutanten unter Verwendung einer großen Anzahl chemischer Verbindungen Informationen über das untersuchte Problem liefern.
Grippe. Behandlung und Prävention.
Influenza ist eine akute Infektionskrankheit der oberen Atemwege. Die an sich gefährliche Grippe verschlimmert den Verlauf anderer chronischer Krankheiten und verursacht schwerwiegende Komplikationen im Herz-Kreislauf- und Zentralnervensystem, den Verdauungsorganen, den Nieren usw. Am gefährlichsten ist die Grippe für Kinder und ältere Menschen. Die Ausbreitungsgeschwindigkeit der Grippe, die Schwere der Erkrankung, die Häufigkeit von Komplikationen und manchmal auch der Tod – all dies macht ihre Prävention besonders wichtig. Menschen, die Sport treiben und turnen, sind deutlich seltener dem Influenzavirus ausgesetzt. Es sind verschiedene Arten von Influenzaviren bekannt – A, B, C usw.; Unter dem Einfluss von Umweltfaktoren kann ihre Zahl zunehmen. Aufgrund der Tatsache, dass die Immunität gegen Influenza kurzfristig und spezifisch ist, ist eine wiederholte Erkrankung in einer Saison möglich. Laut Statistik erkranken jedes Jahr durchschnittlich 20-35 % der Bevölkerung an Grippe.
Die Infektionsquelle ist eine kranke Person; Patienten mit einer milden Form des Virus sind als Überträger des Virus am gefährlichsten, da sie sich nicht rechtzeitig isolieren – sie gehen zur Arbeit, nutzen öffentliche Verkehrsmittel und besuchen Vergnügungsstätten.
Die Infektion wird vom Erkrankten auf den Gesunden durch Tröpfcheninfektion beim Sprechen, Niesen, Husten oder durch Haushaltsgegenstände übertragen.
Die Latenzzeit der Grippe beträgt 1 – 12 Stunden bis 3 Tage. Die Krankheit beginnt akut: starker Temperaturanstieg auf 38-400, Schüttelfrost, Kopfschmerzen, Schmerzen in Knochen und Muskeln, allgemeine Schwäche; Schmerzen und Halsschmerzen, Geschmacks- und Geruchsstörungen treten auf; Nach 12–24 Stunden tritt Nasenausfluss auf.
Die Temperatur hält 1-3 Tage an, manchmal bis zu 6-7 Tage. In der Regel normalisiert sich die Temperatur am Ende der ersten Woche wieder. Bei richtiger Behandlung und Pflege erfolgt die Genesung innerhalb von 7–9 Tagen.
Bei Verdacht auf eine Grippe sollte der Patient isoliert und ins Bett gebracht werden. Dies muss vor Eintreffen des Arztes erfolgen. Da Grippeerreger in der äußeren Umgebung sehr instabil sind und unter dem Einfluss von Sauerstoff und Desinfektionsmitteln leicht zerstört werden, muss der Raum regelmäßig gelüftet werden. Reinigen Sie den Raum mindestens einmal täglich nass mit Bleichmittel, Formaldehyd, Soda, Chloramin und Waschseife.
Der Patient muss individuelle Gerichte haben. Das Geschirr und Teegeschirr des Patienten sollte mit kochendem Wasser und Backpulver gewaschen oder mit einer 5 %igen Chloraminlösung behandelt werden. Eine systematische Desinfektion der Unterwäsche und Bettwäsche des Patienten durch Auskochen in Seifenlösung ist obligatorisch.
Alle vom Arzt verschriebenen Medikamente sollten an einem speziell dafür vorgesehenen Ort aufbewahrt werden. Zusätzlich zu Medikamenten ist es ratsam, in allen Krankheitsphasen viel Flüssigkeit zu sich zu nehmen: Tee mit Honig oder Zitrone, Preiselbeersaft, warme Milch, Obst- und Gemüsesäfte. Das Essen muss kalorienreich sein. Den Anweisungen des Arztes ist unbedingt Folge zu leisten. Selbstmedikation ist inakzeptabel. Medikamente dürfen nicht ohne ärztliche Verschreibung eingenommen werden. Besondere Vorsicht ist bei Antibiotika und Sulfonamiden geboten – sie wirken nicht auf das Influenzavirus und können bei unerlaubter Einnahme und falscher Dosierung allergische Reaktionen hervorrufen. Was Sie aber schmerzfrei anwenden können, sind Fußbäder, Senfpflaster, Zwiebeln, Knoblauch. Über die Lunge abgegeben, steigern die in Zwiebeln und Knoblauch enthaltenen ätherischen Öle die Schleimsekretion und erleichtern so das Abhusten bei Atemwegserkrankungen.
Das Gesundheitswesen verfügt außerdem über eine Reihe spezifischer Anti-Influenza-Mittel, zu denen vor allem ein Lebendimpfstoff und ein spezielles Serum mit schützenden Proteinen gehören. Die in der Praxis eingeführten Medikamente sind Interferon und Oxolinsalbe.
Abhärtung, ausgewogene Ernährung, frische Luft und die rechtzeitige Behandlung chronischer Krankheiten helfen Ihnen, Erkältungen, insbesondere Grippe, vorzubeugen.
Es wird angenommen, dass Viren (von lat. Virus- Gift) - etwas Böses, das nur Ärger bringt. Aber das ist ein schwerwiegender Fehler. Viren sind die wichtigsten Schöpfer der belebten Natur und die Motoren ihrer Evolution.
Eines der Hauptargumente gegen die Hypothese, dass Viren aus Zellen „entkommen“ seien, ist die Tatsache, dass virale genetische Systeme viel vielfältiger sind als zelluläre. Wie bekannt ist, verfügen zelluläre Organismen ausschließlich über doppelsträngige – lineare oder zirkuläre – DNA-Genome. Und das Genom des Virus kann sowohl durch einzel- als auch doppelsträngige RNA- oder DNA-Moleküle, linear oder zirkulär, dargestellt werden. Es gibt auch Systeme, die die umgekehrte Transkription nutzen. So wird bei Retroviren (z. B. einigen Onkoviren, HIV) und Pararetroviren (Hepatitis-B-Viren, Blumenkohlmosaik usw.) eine der genomischen DNA-Ketten auf einer RNA-Matrize synthetisiert. Im Gegensatz zu zellulären Organismen nutzen Viren alle theoretisch möglichen Möglichkeiten, genetische Informationen zu speichern und auszudrücken.
Das zweite wichtige Argument dagegen, Viren als aus Zellen stammend zu betrachten, ist, dass es viele virale Gene gibt, die in zellulären Organismen nicht existieren. Zelluläre Organismen haben sich nicht nur aus Viren entwickelt, sondern haben auch einen erheblichen Teil ihres genetischen Materials von ihnen geerbt (und erben dies auch weiterhin). Von besonderem Interesse sind in diesem Zusammenhang endogene Viren (Teile des Genoms von RNA- oder DNA-Viren, die in das Genom einer Zelle integriert sind), wobei von Retroviren abgeleitete Gene überwiegen. Es wird angenommen, dass Säugetiere mehr als die Hälfte ihres Genoms von Viren und ihren nächsten Verwandten geerbt haben – „egoistischen“ genetischen Elementen, zum Beispiel Plasmiden und Transposons. Somit sind Viren Miteltern des Menschen. Oft sind die Gensequenzen endogener Viren, die im menschlichen Genom in großer Zahl vorkommen, verändert und kodieren nicht mehr für Proteine. Es gibt gute Gründe zu der Annahme, dass solche Sequenzen an der Regulierung zellulärer Gene beteiligt sind, obwohl ihre spezifischen biologischen Funktionen oft unbekannt sind. Allerdings wissen wir etwas Wichtiges: Beispielsweise ist das Protein Syncytin, das vom Hüllgen eines endogenen Retrovirus kodiert wird, für die Zellfusion bei der Plazentabildung notwendig. Das bedeutet, dass weder Menschen noch Plazentatiere ohne dieses endogene Virus geboren werden könnten. Es gibt noch ein weiteres wichtiges Beispiel. Es stellte sich heraus, dass ein Bestandteil des Genoms eines der endogenen Viren die Expression der Prolin-Dehydrogenase in einigen Regionen des Zentralnervensystems steuert. Dieses Enzym könnte eine wichtige Rolle bei der Entwicklung des menschlichen Gehirns gespielt haben. Ist die Expression dieses Enzyms durch Mutationen gestört, kommt es zu psychischen Erkrankungen bis hin zur Schizophrenie. Auch bei der horizontalen Übertragung zellulärer Gene – von einem Organismus auf einen anderen – spielen Viren und ihre Verwandten eine wichtige Rolle.
Doch trotz ihrer Schlüsselrolle in der Evolution sind Viren vor allem als Krankheitserreger von Menschen, Tieren und Pflanzen bekannt (übrigens wurden sie deshalb erstmals entdeckt). Und dann werden wir über die Natur der viralen Pathogenität sprechen. Viren (insbesondere eukaryotische Viren) haben kein besonderes „Verlangen“, dem Wirt Schaden zuzufügen, geschweige denn ihn zu töten. Und in vielen Fällen koexistieren Viren recht friedlich und freundlich mit Zellen. Warum sind so viele Viren so schädlich? Die übliche Erklärung ist, dass die Pathologie einer infizierten Zelle durch die „Plünderung“ ihrer (materiellen und strukturellen) Ressourcen verursacht wird, die das Virus für seine eigenen Fortpflanzungsbedürfnisse verwendet. Der größte Schaden kann jedoch durch unüberlegte Schutzmaßnahmen von Wirten und die Schutzaktivität von Viren entstehen, die nicht direkt mit ihrer Reproduktion zusammenhängt.
Schutz- und Gegenverteidigungsmechanismen
Was sind die wichtigsten Abwehrmechanismen einer infizierten Zelle? Dies sind Bestandteile der angeborenen Immunität: Abbau der RNA (sowohl viral als auch zellulär), Hemmung der Proteinsynthese (sowohl viral als auch zellulär), Selbstzerstörung (Apoptose und andere Arten des programmierten Todes) und schließlich Entzündung. Tatsächlich haben viele Viren auf diese Weise ihre Existenz entdeckt – aufgrund der von ihnen verursachten Entzündungen (Enzephalitis, Lungenentzündung usw.). Die Zelle bekämpft das Virus, indem sie ihren eigenen Stoffwechsel und/oder ihre eigene Struktur stört, und ihre Abwehrmechanismen sind in der Regel selbstschädigend. Man könnte sagen, dass eine Person, die an Polio stirbt (und das sind weniger als 1 %), sich im Kampf gegen die Infektion das Leben genommen hat.
Als Reaktion auf zelluläre Abwehrkräfte entwickeln sich Viren weiter, um Abwehrkräfte zu entwickeln, und es kommt zu einem Wettrüsten zwischen dem Virus und der Zelle. Diese Medikamente richten sich vor allem gegen die allgemeinen Stoffwechselprozesse, die den Abwehrreaktionen der Zelle zugrunde liegen. Dabei handelt es sich wiederum um eine Hemmung der Synthese zellulärer RNA und Proteine, eine Störung der intrazellulären Infrastruktur und des Zelltransports, eine Unterdrückung oder umgekehrt die Auslösung von Apoptose und anderen Mechanismen, die den programmierten Zelltod verursachen. Somit ähnelt die Abwehrstrategie eines Virus in vielerlei Hinsicht dem Abwehrverhalten einer Zelle. Im übertragenen Sinne nutzen Wrestler die gleichen Techniken und erreichen die gleichen Ziele. Beispielsweise verwendet eine Zelle, die die Synthese viraler Proteine unterdrückt, Interferon, und um dessen Bildung zu hemmen, hemmt das Virus wiederum die Proteinsynthese in der Zelle. Abhängig von den Umständen profitiert die eine oder andere Seite. Es stellt sich heraus, dass der Hauptbeitrag zur Pathologie nicht die Reproduktion des Virus als solches ist, sondern die Konfrontation zwischen zellulärer Abwehr und viraler Abwehr. In der Pflanzenpathologie gibt es seit langem das Konzept der „Toleranz“: Ein pathogenes Virus kann sich in einer infizierten Pflanze aktiv vermehren, ohne schmerzhafte Symptome zu verursachen.
Im Folgenden werden wir hauptsächlich über RNA-Viren sprechen (dies ist ein einfacheres Beispiel). Wie verrät ein RNA-Virus seine Anwesenheit, sobald es in eine Zelle gelangt? Und woher weiß eine Zelle, dass ein Virus in sie eingedrungen ist? Das Hauptmerkmal, aufgrund dessen die Zelle dies „versteht“, ist virale doppelsträngige RNA, die grundsätzlich in einer nicht infizierten Zelle gebildet werden kann, jedoch nicht in solchen Mengen und an solchen Orten. In einigen Fällen erkennt die Zelle auch virale einzelsträngige RNA und manchmal (viel seltener) virale Proteine. Es ist wichtig, dass die Erkennung viraler RNA unspezifisch ist: Nachdem die Zelle doppelsträngige RNA „gespürt“ hat, „glaubt“ sie möglicherweise, dass ein Virus in sie eingedrungen ist, weiß aber nicht, welches. RNA wird von zwei Arten von Sensoren erfasst: Toll-like (aus dem Englischen). mautartig und von ihm. Maut- bemerkenswert) Rezeptoren und spezialisierte RNA-Helikasen. Sie beinhalten eine Reihe von Schutzmechanismen auf der Transkriptionsebene, darunter die Bildung von Interferon. Darüber hinaus werden virale RNAs von den „Ausführern“ erkannt – der doppelsträngigen RNA-abhängigen Proteinkinase PKR, die einige Translationsinitiationsfaktoren phosphoryliert und dadurch die Proteinsynthese hemmt; Oligoadenylatsynthetase (OAS), die RNase L aktiviert, die RNA spaltet; RNA-Interferenzsystem, das zum Abbau der RNA und zur Störung ihrer Translation führt.
Da das Virus als etwas Unspezifisches erkannt wird, kann die Zelle seine „Absichten“ nicht kennen. Und im Allgemeinen wäre es unmöglich, ein individuelles angeborenes Abwehrsystem für jeden möglichen Virus zu entwickeln. Das bedeutet, dass die Zelle das Virus nur mit Standardtechniken bekämpfen kann. Und deshalb stehen seine Abwehrmaßnahmen oft in keinem Verhältnis zur bestehenden Bedrohung. Wenn aber die Abwehrreaktionen der Zelle so unspezifisch sind, warum verursachen dann unterschiedliche Viren immer noch unterschiedliche Krankheiten? Erstens kann jedes Virus nur einen bestimmten Zelltyp in einem bestimmten Organismus infizieren. Dies liegt daran, dass es, um in die Zelle einzudringen, mit dafür „geeigneten“ zellulären Rezeptoren interagieren muss. Darüber hinaus ist für die Vermehrung von Viren eine bestimmte intrazelluläre Umgebung erforderlich (häufig werden spezifische zelluläre Proteine benötigt). Zweitens: Während die Abwehrreaktionen der Zelle Standard sind, sind die Abwehrreaktionen des Virus weitgehend individuell, obwohl sie sich gegen standardmäßige zelluläre Mechanismen richten.
In Pflanzen spielt die RNA-Interferenz als antiviraler Mechanismus eine sehr wichtige Rolle. Doppelsträngige RNA wird aus viraler RNA gebildet (ein wichtiger Faktor, durch den die Zelle von der Anwesenheit eines Virus erfährt). Unter Beteiligung von Komponenten des RNA-Interferenzsystems – dem Dicer-Enzym, das diese doppelsträngige RNA in Fragmente von 21–25 Nukleotidpaaren Länge schneidet, und dann des RNA-Protein-Komplexes RISC – entstehen schließlich einzelsträngige kurze RNA-Fragmente gebildet. Durch Hybridisierung mit viraler RNA bewirken sie entweder deren Abbau oder die Hemmung ihrer Translation. Dieser Schutzmechanismus ist wirksam, kann jedoch die Zelle selbst schädigen, wie am Beispiel der Viroide deutlich zu sehen ist. Dabei handelt es sich um Pflanzenpathogene, kurze (mehrere hundert Nukleotide) kreisförmige einzelsträngige RNA-Moleküle, die nicht mit einer Proteinhülle bedeckt sind. Viroide kodieren nicht für Proteine, können aber bei einer infizierten Pflanze schwere Symptome hervorrufen. Dies geschieht, weil die Zelle sich selbst schützt. Die resultierende viroide doppelsträngige RNA wird allen Komponenten des RNA-Interferenzsystems ausgesetzt, was zur Bildung einzelsträngiger RNA-Fragmente führt, die nicht mit viraler RNA, sondern mit zellulärer RNA hybridisieren. Dies führt zu dessen Abbau und der Entwicklung von Krankheitssymptomen. Viele Pflanzenviren kodieren jedoch für eine Vielzahl von Proteinen, die die RNA-Interferenz stören ( virale Suppressoren der RNA-Stummschaltung – VSR). Sie hemmen entweder die Erkennung und Spaltung viraler RNAs oder unterdrücken die Bildung und Funktion des RISC-Komplexes. Daher können diese VSR-Proteine physiologisch wichtige (nicht-virale) RNA-Interferenzmechanismen stören und pathologische Symptome verursachen.
Sicherheitsproteine
Die Abwehr von Viren, insbesondere Picornaviren, kleinen RNA-haltigen Krankheitserregern, hängt maßgeblich von der Arbeit solcher Proteine ab. Zu dieser großen Gruppe gehören insbesondere die Erreger von Poliomyelitis, Hepatitis A, Maul- und Klauenseuche etc. Die Besonderheit dieser Viren besteht darin, dass ihre Proteine bis auf seltene Ausnahmen in Form eines einzigen Polyproteins synthetisiert werden welche einzelnen reifen Proteine dann gebildet werden. Unter ihnen lassen sich drei Gruppen unterscheiden. Das erste besteht aus Schlüsselproteinen – lebenswichtig, mit festen Funktionen, die direkt die Reproduktion des Virus gewährleisten: RNA-abhängige RNA-Polymerasen, die für die Replikation des viralen Genoms notwendig sind; Kapsidproteine, die die Proteinhülle des Virus bilden; Proteasen, die am Prozess der Umwandlung von Polyproteinen in reife Proteine beteiligt sind; VPg-Protein ( virales Proteingenom verknüpft- ein mit dem Genom verbundenes virales Protein, das als Primer für die Synthese von RNA-Molekülen dient; Helicase ist ein sehr wertvolles Enzym, das alle Picornaviren besitzen, dessen Rolle jedoch nicht ganz klar ist. Zur zweiten Gruppe gehören auch lebenswichtige Proteine, aber solche, die „Hilfsarbeit“ leisten – die hydrophoben „Leitproteine“ 2B und 3A. Sie leiten wichtige Proteine an ihren Bestimmungsort und tragen dazu bei, eine optimale intrazelluläre Umgebung für die Virusvermehrung zu schaffen. Zur dritten Gruppe gehören das vor 30 Jahren in unserem Labor entdeckte Leitprotein L und Protein 2A; wir nannten sie „Sicherheits“-Proteine ( Sicherheit- Sicherheit). Dabei handelt es sich um eine spezielle Abwehrwaffe gegen Picornaviren. Im Allgemeinen können alle drei dieser Proteinklassen die Abwehrmechanismen der Zelle bekämpfen. Aber Schlüsselproteine und Leitproteine erledigen diese Aufgabe nebenbei, weil sie andere wichtige Aufgaben haben, denen ihre Struktur und Funktion gerecht werden muss. Folglich sind ihre Verteidigungsfähigkeiten durch die Notwendigkeit, grundlegende Arbeiten auszuführen, eingeschränkt. Aber „Sicherheits“-Eichhörnchen arbeiten Vollzeit in ihrem Fachgebiet – die Evolution hat sie speziell für „Sicherheit“ „eingestellt“ (später „lernten“ einige von ihnen, etwas anderes zu tun). Sie können über jede zur Erfüllung ihrer Aufgaben erforderliche Struktur verfügen.
Eine der wichtigsten Funktionen von Sicherheitsproteinen besteht darin, dass sie das Schicksal einer infizierten Zelle bestimmen. Es gibt viele verschiedene Möglichkeiten für seinen Tod, aber die beiden wichtigsten und bekanntesten Mechanismen sind Nekrose und Apoptose, die sich in ihren morphologischen und biochemischen Eigenschaften unterscheiden. Bei der Nekrose wird die Zelle aufgelöst und ihr Inhalt in den Interzellularraum ausgeschüttet. Während der Apoptose bilden sich deutlich sichtbare Vorsprünge auf ihrer Oberfläche, ihre DNA wird zu nukleosomalen Fragmenten abgebaut und schließlich wird die Zelle in einzelne apoptotische Körper fragmentiert, die auf die Plasmamembran begrenzt sind. Es ist sehr wichtig, wie genau die Zelle stirbt. Bei der Nekrose entwickelt sich eine schützende Entzündung, das Virus verlässt jedoch die Zelle und breitet sich aus. Bei der Apoptose ist die Ausbreitung des Virus begrenzt und es kommt in der Regel zu keiner Entzündungsreaktion. Der Tod einer infizierten Zelle ist in der Regel ein Akt der Selbstaufopferung, der die Vermehrung des Virus einschränkt.
Wir fanden heraus, dass eine Infektion mit Picornaviren, insbesondere dem Poliovirus (Poliovirus), ein zellapoptotisches Programm auslöst. Dies geschieht auf einem der klassischen Wege, wenn Cytochrom aus Mitochondrien freigesetzt wird C und eine Kaskade proteolytischer Enzyme, Caspasen, wird aktiviert. Andererseits stellte sich heraus, dass Viren über einen antiapoptotischen Mechanismus verfügen – die Fähigkeit, die apoptotische Reaktion der Zelle zu unterdrücken. So sterben HeLa-Zellen, die mit Poliovirus oder Enzephalomyokarditisvirus (auch Picornavirus) infiziert sind, mit Anzeichen einer Nekrose ab. Wenn Sie jedoch die antiapoptotische „Waffe“ ausschalten (die Synthese viraler Proteine unterdrücken), stirbt die Zelle an Apoptose (Selbstaufopferung). Bei beiden Viren dienen „Sicherheits“-Proteine als solche Waffen. Beim Enzephalomyokarditisvirus übernimmt diese Rolle jedoch das L-Protein, beim Poliovirus ist es das 2A-Protein. Das Leader-Protein hat keine enzymatische Aktivität, während das 2A-Protein eine Protease ist. Sie haben weder strukturell noch biochemisch etwas gemeinsam, beide haben jedoch antiapoptotische Wirkungen, die auf unterschiedlichen molekularen Mechanismen beruhen.

Ein weiterer Schutzmechanismus der Sicherheitsproteine des Picornavirens ist die Störung des nuklear-zytoplasmatischen Transports [10–12]. Wir haben gezeigt, dass bei einer Infektion mit diesen Viren die Permeabilität der Kernmembran zunimmt und der aktive Austausch von Makromolekülen zwischen Zytoplasma und Kern gestört wird. Und wenn die Zellstruktur geschädigt ist, kann sie ihre Regulationsmechanismen zur Bekämpfung des Virus nicht aktivieren. Beim Poliovirus stört das Sicherheitsprotein 2A den nuklear-zytoplasmatischen Transport durch Hydrolyse von Nukleoporinen, Bestandteilen der Kernporen. Und das Leitprotein des Enzephalomyokarditis-Virus wirkt – es beeinflusst die zelluläre Phosphorylierungskaskade von Nukleoporinen [,].
Die Anti-Schutzfunktion von „Sicherheitsproteinen“ kann sich auch auf andere Weise manifestieren. So hemmen L-Proteine von Kardioviren (einschließlich Enzephalomyokarditis-Virus) und 2A-Proteine von Enteroviren (einschließlich Poliovirus) die Bildung von Interferon. Und seine Wirkung wird durch das L-Protein des Maul- und Klauenseuchevirus und das 2A-Protein des Poliovirus gehemmt. Allerdings sind die „Sicherheits“-Proteine der Picornaviren nicht lebenswichtig. Es können beide Wächter entfernt oder große Deletionen in ihnen verursacht werden (wie im Fall des L-Proteins von Kardioviren, der 2A-Proteine des Hepatitis-A-Virus und von Kardioviren), ohne dass das Virus lebensfähig gemacht wird.
Gegenseitige Abrüstung

Welche Folgen hat die Inaktivierung viraler „Sicherheitsproteine“ für die Zelle? Einerseits wird die Empfindlichkeit von Viren gegenüber den Schutzmechanismen der angeborenen zellulären Immunität zunehmen. Andererseits wird aber auch seine selbstverletzende, suizidale Aktivität zunehmen. Was passiert, wenn man gleichzeitig die Abwehrmechanismen der Zelle und des Virus ausschaltet? Wir haben diese Situation am Beispiel der Interaktion zwischen Mengovirus (einem Stamm des Enzephalomyokarditis-Virus) und HeLa-Zellen untersucht. Bei einer Infektion mit einem Wildtyp-Virus sterben sie schnell an Nekrose. Und wenn das Virus teilweise entwaffnet wird (das Leitprotein wird inaktiviert), leben HeLa-Zellen etwas länger und sterben nicht an Nekrose, sondern an Apoptose. Wenn die gegenseitige Abwehr verringert ist (die Apoptose wird in der Zelle durch eine chemische Verbindung, die Caspasen hemmt, ausgeschaltet und das Leitprotein des Virus inaktiviert), fühlen sich die Zellen selbst nach einer doppelt so langen Zeitspanne deutlich besser als diejenigen, die dies nicht waren entwaffnet. Und die Vermehrung des Virus (sowohl Dynamik als auch Ernte) verlief genauso, unabhängig davon, ob nur seine Abwehrkräfte ausgeschaltet wurden oder ob gleichzeitig auch die zellulären Abwehrkräfte entfernt wurden. Es zeigt sich, dass sich in Zellen, die noch keine ernsthafte pathologische Schädigung (den sogenannten zytopathischen Effekt) aufweisen, bereits viele Viruspartikel bilden können. Somit ist eine Zellschädigung nicht notwendig, damit sich das Virus vermehren kann. Daher kann eine wirksame Strategie zur antiviralen Therapie zur Linderung der Krankheitssymptome die gleichzeitige Unterdrückung sowohl der viralen als auch der zellulären Abwehr sein.
Programmierter Tod
Diese Reihe unserer Experimente bot auch die Gelegenheit, tiefer in die Natur des durch das Virus verursachten nekrotischen Todes einzudringen. Was ist das – die Tötung einer Zelle durch einen Virus oder ihr Selbstmord (Selbstaufopferung), wenn sie entscheidet, dass es aus Gründen des Gemeinwohls sinnvoller ist zu sterben? Anfällig für nekrotische Läsionen sind:
- Plasmamembran (ihre Durchlässigkeit nimmt zu, es bilden sich „Blasen“),
- Zytoplasma (Mikrotubuli und Mikrofilamente verändern sich),
- Zellkern (schrumpft, verformt sich, Chromatin kondensiert),
- Stoffwechselaktivität (NADH-abhängige Reduktionsreaktionen, Lebensfähigkeitsänderung).
Wenn die Apoptose ausgeschaltet wird (die Zugabe eines chemischen Caspase-Inhibitors), hängen verschiedene nekrotische Veränderungen davon ab, ob das virale Leitprotein funktioniert oder nicht. Wenn es beispielsweise inaktiviert wird, ändert sich die Membranpermeabilität der Zelle nicht, es treten keine „Blasen“ auf, die mit einem Ungleichgewicht im osmotischen Gleichgewicht verbunden sind, und eine Reihe anderer nekrotischer Läsionen treten nicht auf. Eine mögliche Erklärung für diesen Effekt ist, dass das L-Protein auf mehrere Ziele in verschiedenen Zellkompartimenten einwirkt. Da das Protein jedoch klein ist und keine enzymatische Aktivität aufweist, ist es wahrscheinlicher, dass seine direkten Ziele viel kleiner sind. Wir gehen davon aus, dass das Leitprotein auf ein oder mehrere wichtige zelluläre Elemente einwirkt, die das Schicksal der Zelle steuern, und dadurch ihr nekrotisches Programm in Gang setzt, das für die meisten der aufgeführten pathologischen Veränderungen verantwortlich ist. Folglich ist es nicht das Virus, das die Zelle nekrotisch tötet, sondern die Zelle selbst begeht Selbstmord (führt einen Akt der Selbstaufopferung aus). Diese Sichtweise steht im Einklang mit neuen Vorstellungen, wonach es neben der Apoptose eine Reihe weiterer physiologisch wichtiger Arten des programmierten (im Zellgenom kodierten) Zelltods gibt, darunter die der Nekrose ähnliche Nekroptose.
Daher kann sich die Selbstaufopferung der Zellen während einer Virusinfektion als Nekroptose oder Apoptose äußern. Nekroptose kann nicht nur bei Picornaviren eine Schutzreaktion einer Zelle auf eine Virusinfektion sein. Welcher Mechanismus für das Virus vorteilhafter ist, hängt von den Bedingungen ab. Wir sehen, dass sich seine antiprotektive Wirkung in Form einer „Umleitung“ von im Zellgenom kodierten Mechanismen manifestieren kann. Dies ist eine wichtige (wenn auch nicht die einzige) Abwehrmethode und einer der Hauptmechanismen der Pathogenität von Viren. Virusinduzierte Apoptose- und Nekroseprogramme konkurrieren miteinander. Wir haben gezeigt, dass bei einer Infektion von HeLa-Zellen mit dem Poliovirus zunächst die Apoptose aktiviert, dann unterdrückt und der nekrotische Weg in Gang gesetzt wird. Somit aktiviert die Infektion einer Zelle mit einem Virus eine Reihe von Schutzmaßnahmen in ihr, darunter zwei selbstmörderische Mechanismen des programmierten Todes – apoptotisch und nekrotisch. Und dann kommt es zu einem Wettbewerb zwischen diesen Pfaden: Die Unterdrückung eines von ihnen aktiviert den anderen und umgekehrt. Und all dies wird durch zelluläre Proteine, virale Proteine (hauptsächlich „Sicherheits“-Proteine) sowie externe Faktoren reguliert.
Wettrüsten
Da Zellen über Abwehrmechanismen verfügen und Viren über Abwehrmechanismen verfügen, kommt es zwischen ihnen natürlich zu einem Wettrüsten. Die nichtkonservative Natur von Sicherheitsproteinen legt nahe, dass sie dazu geeignet sind, den Abwehrmechanismen eines bestimmten Wirts entgegenzuwirken. Und daher kann seine Veränderung mit einem Verlust der „Sicherheits“-Proteinfunktion und infolgedessen einer Zunahme der Abwehrreaktionen des Wirts einhergehen. Dies kann die besondere Pathogenität des „Neuen“ erklären ( neu entstehend- neu auftretende) Viren. Somit handelt es sich beim Influenzavirus um ein wenig pathogenes, nahezu harmloses Darmvirus von Wildvögeln. Wenn eine Person infiziert wird, kann es zur Spanischen Grippe, Vogelgrippe oder Schweinegrippe kommen. Das SARS-Virus ist für Fledermäuse relativ ungefährlich, beim Menschen verursacht es jedoch ein schweres akutes Atemwegssyndrom, das mit einer hohen Sterblichkeit einhergeht. Schließlich ist HIV (genauer gesagt sein Vorfahre) für Affen praktisch harmlos, beim Menschen verursacht es jedoch AIDS. Es ist sehr wichtig, dass diese Viren bei der Übertragung auf einen neuen Wirt keine neuen Pathogenitätsfaktoren entwickeln (sie erwerben lediglich durch mehrere Mutationen, die das Eindringen in die Zelle gewährleisten, die Fähigkeit, Menschen zu infizieren). Ein weiterer möglicher Mechanismus für das Ungleichgewicht zwischen Virus und Wirt und die Entstehung neuer Krankheitserreger könnte eine Veränderung der viralen Abwehrwaffen sein, beispielsweise der Verlust eines alten oder der Erwerb eines neuen „Sicherheits“-Proteins.
Eine langfristige Koevolution von Wirt und Virus sollte jedoch zu einer Verringerung der Pathogenität des Virus führen (für beide Seiten vorteilhafte Abrüstung). Das klassische Beispiel ist das Myxoma-/Fibromvirus. Mitte des 19. Jahrhunderts. Europäische Kaninchen wurden nach Australien gebracht, sie vermehrten sich schnell und wurden zu einer ernsthaften Bedrohung für die Landwirtschaft. 100 Jahre später begann man, das pathogene Fibrom-/Myxomavirus (aus der Familie der Pockenviren, zu der auch das Pockenvirus gehört) zur Kontrolle ihrer Population einzusetzen. Verschiedene Kaninchen reagieren unterschiedlich auf dieses Virus. Bei brasilianischen Kaninchen kommt es drei Wochen nach der Infektion zu einem gutartigen Tumor – einem Fibrom (lokal begrenzter Knoten auf der Haut). Aber europäische Kaninchen, die empfindlich auf dieses Virus reagieren, entwickeln innerhalb von 10 Tagen nach der Infektion eine generalisierte tödliche Krankheit.
Das in Australien eingeschleppte Virus verursachte durch Mücken übertragene Sommerepidemien, bei denen mehr als 99 % der infizierten Kaninchen in weniger als zwei Wochen starben. Weniger virulente Varianten des Virus überlebten den Winter eher, was zur Selektion geschwächter (attenuierter) Stämme führte. Und nach etwa 10 Jahren halbierte sich die Sterblichkeitsrate europäischer Kaninchen aufgrund des entwickelten Virus. Gleichzeitig wurden resistente Kaninchen selektiert: Ihre Sterblichkeit durch das ursprüngliche Virus verringerte sich um etwa das Vierfache. In nur einem Jahrzehnt (evolutionär gesehen ein vernachlässigbarer Zeitraum) hat sich die Beziehung zwischen Krankheitserreger und Wirt etwa um das Zehnfache verbessert. Dies ist natürlich ein etwas vereinfachtes Schema, da das Wettrüsten nicht aufhört: Als Reaktion auf eine erhöhte Resistenz bei Kaninchen kann auch die Virulenz des Virus zunehmen. Dies ist jedoch ein eindrucksvolles Beispiel für die Rolle der Interaktion von Viren und Zellorganismen bei der Evolution beider. Viren und Zellen „lehren“ sich gegenseitig und das erworbene „Wissen“ wird vererbt. Im Jahr 2013 veröffentlichten zwei Absolventen der Abteilung für Virologie der Moskauer Staatlichen Universität, Evgeny Kunin und Valeryan Dolya, einen Artikel über die „virozentrische“ Sichtweise der Evolution, wonach die Resistenz und Zusammenarbeit von Viren und Zellorganismen der Hauptfaktor sind ihre Entwicklung.
Meine Geschichte erschöpft das Thema nicht: Es ist noch viel mehr über die Pathogenität von Viren bekannt. Vieles von dem, was wir heute wissen, wurde in den letzten Jahren gelernt, und es gibt allen Grund, mit neuen Überraschungen zu rechnen. Wir können und sollten Viren für schwere Krankheiten verantwortlich machen und es ist notwendig, sie zu bekämpfen, aber wir sollten den Viren für die Existenz und Vielfalt der belebten Natur, einschließlich der Existenz des Menschen, dankbar sein.
Der Autor dankt seinen Kollegen in der wissenschaftlichen Zusammenarbeit – den Mitarbeitern des nach ihm benannten Instituts für Poliomyelitis und virale Enzephalitis. M. P. Chumakov RAMS, Moskauer Staatliche Universität. M. V. Lomonosov, Institut für Protein RAS (Pushchino, Region Moskau), Universität Basel (Schweiz), University of Wisconsin (USA), Universität Nijmegen. Radboda (Niederlande).
Der Artikel basiert auf einem Vortrag an der Schule „Moderne Biologie und Biotechnologien der Zukunft“ (Zvenigorod, 26. Januar – 1. Februar 2014).
. Belov G. A., Lidsky P. V., Mikitas O. V. et al. Bidirektionaler Anstieg der Permeabilität der Kernhülle bei Poliovirus-Infektion und begleitenden Veränderungen der Kernporen // J. Virol. 2004. V. 78. S. 10166–10177. doi:10.1128/JVI.78.18.10166-10177.2004. Lidsky P. V., Hato S., Bardina M. V. et al. Durch Kardioviren induzierte nukleozytoplasmatische Verkehrsstörung // J. Virol. 2006. V. 80. S. 2705–2717. doi:10.1128/JVI.80.6.2705-2717.2006
. Bardina M. V., Lidsky P. V., Sheval E. V. et al. Mengovirus-induzierte Neuordnung des Kernporenkomplexes: Kaperung der zellulären Phosphorylierungsmaschinerie // J. Virol. 2009. V. 83. S. 3150–3161. doi:10.1128/JVI.01456-08
. Mikitas O. V., Ivin Y. Y., Golyshev S. A. et al. Unterdrückung von Verletzungen durch ein lytisches RNA-Virus (Mengovirus) und deren Entkopplung von der Virusvermehrung durch gegenseitige Zell-/Virusentwaffnung // J. Virol. 2012. V. 86. S. 5574–5583. doi:10.1128/JVI.07214-11
. Agol V. I., Belov G. A., Bienz K. et al. Konkurrierende Todesprogramme in Poliovirus-infizierten Zellen: Verpflichtungswechsel mitten im Infektionszyklus // J. Virol. 2000. V. 74. S. 5534–5541. doi:10.1128/JVI.74.12.5534-5541.2000
. Agol V. I. Zytopathische Wirkungen: virusmodulierte Manifestationen der angeborenen Immunität? // Trends Mikrobiol. 2012. V. 20. S. 570–576. doi:10.1016/j.tim.2012.09.003
. Koonin E. V., Dolja V. V. Eine virozentrische Perspektive auf die Evolution des Lebens // Curr. Meinung. Virol. 2013. V. 3. S. 546–557. doi:10.1016/j.coviro.2013.06.008